A look at Endosymbiosis- a crazy idea that worked
While researching for the latest chapter in my Prometheus fanfic I stumbled (well, stumbled again) on the theory of endosymbiosis. Which is that, sometimes, a collaboration between two or more organisms, who may even have been in a predatory race at some point, results in them permanently merging into one single individual. In particular, the structure of the eukariotic cell, which is the type of cell that animals and multicellular plants are made of, appears to be the result of several endosymbiotic events.
Since Prometheus the film deals with lofty things as the origin of life and whatnot, I thought endosymbiosis is a worthy topic to try and reference, however oblique and distant the reference ends up in the text. For one, it neatly contrasts with the view of life as eternal, deadly competition. That's a true view, by the way; the circle of life isn't nearly as nice as new age sentiment makes it to be. Life is vicious, cruel, short-sighted and selfish. Paradoxically, this blind idiot monster sometimes produces incredibly long-lasting partnerships, and trying to explain how that may happen is the point of this post. (In the fic, the endosymbiosis is engineered by intelligent agents, the main characters. For the real life endosymbiotic partnerships, to the best available evidence, there was no such thing.)
Another reason why endosymbiosis is interesting, to me, is that it's a genuinely cool, genuinely mad idea that, unlike most mad ideas in science, withstood the test of evidence and came out the stronger for it. You might think, if you went by the crap that passes for pop-sci these days, that crazy and worthy ideas happen all the time: relativity, quantum mechanics, quantum electrodynamics, chaos theory and so on. False. Those first three were built on a solid foundation of experiments that older theories couldn't explain. The last (chaos theory) was simply the natural consequence of classical mechanics as revealed by the high powered mathematics emerging at the end of the nineteenth century. All of that may be fodder for other posts though.
No, a better example of a similarly crazy and worthy idea is continental drift. When Alfred Wegener proposed that continents slowly move across the surface of the Earth, he really had nothing but a hunch based on the similarity of the coasts of South America and Africa. When Konstantin Mereschkowski and Andreas Schimper proposed that chloroplasts, tiny structures in plant cells that are responsible for photosynthesis, were originally independent bacteria, they really only had a hunch based on the fact that chloroplasts kinda looked like bacteria.
That was scant evidence, at best. It so happens however that in these two cases the hunches turned out to be true. To the best of our knowledge today, chloroplasts indeed descend from cyanobacteria that were, in some distant past, independent of the plant cell. One aspect of that proof is that they have their own DNA with great similarities to that of cyanobacteria. Mitochondria, structures in both plant and animal cells that produce ATP to fuel the cells' other activities, also used to be independent bacteria eons ago. They carry their own DNA too, and since only the egg cell transports mitochondria into a foetus, using mitochondrial DNA one can trace the maternal lineage of somebody.
Now, a disclaimer. I'm not a biologist. I will try to give an honest, clear and concise summary of what I understand about evolution and endosymbiosis though, even if all I can produce is, in fairness, a just-so story. Still, I think I'm not too far off in how I imagine the thing to work. So onwards, beginning with a brief summary on evolution.

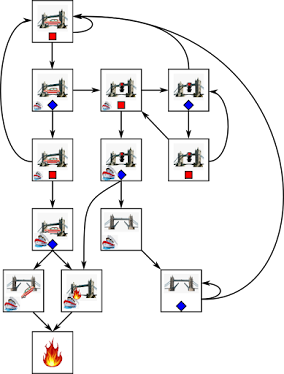
First, mutations. In the figure I tried to represent how mutations spread in a population of bacteria that reproduce asexually. Each bacteria will, if it lives long enough, split itself into two smaller bacteria at some point, which will grow to normal size using nutrients from their surroundings. Initially there's one 'blue' bacteria, and I'll often refer to all of its descendents together as a 'population'.
Even if at some point not all descendents are alike. In the figure, the original bacteria splits into two, and these two split again to make four, but a mutation occurs and now one of the bacteria is 'red'. Next generation, the blue bacteria split into more blue bacteria, while the red is now the ancestor of a small population of red bacteria. Both versions, 'blue' and 'red' may coexist in the population of descendants of the original blue.
Sometimes a mutation is so harmful that it prevents the organism it appears in from reproducing. Such a mutation is deleterious, like the 'green' in the figure. Since green doesn't live long enough to get descendants, that mutation disappears.
Most mutations are neutral. In the figure, 'red' and 'blue' are more or less doing equally well. Context, that is, the environment that organisms live in, is what will select among mutations. Some mutations will make the organism that carries them better able to get descendants in the environment it lives in. In the figure, 'purple' is especially prolific, and will tend to dominate the population of descendants of the original blue.
Note, this doesn't necessarily mean that blue and red disappear. Unless there's something in the environment that actively kills them off, they'll stick around. In lesser numbers than purple, true. But evolution is NOT about being the most efficient, nor the most prolific. As long as you can get a future generation, you're doing well enough (of course, being prolific and efficient helps).
With that basis set up, let's tackle endosymbiosis. For this, imagine the following scenario. Suppose there's a population of single celled animals like amoebas or something similar, merrily going about their single celled lives. These will be the 'hosts'. Now, there are also, floating around, some parasitic bacteria that have the nasty habit of infiltrating inside a host cell, feasting on its contents and multiplying prodigiously, killing it in the process. These will be the 'parasites'.

So the parasites cause a deadly disease in a host, if they infect it. So far, so fine. Evolution in a typical host-parasite competition. Because, suppose one of the host cells suffers a mutation that makes it immune to the parasite infection. They can't get in or are killed off if they do, whatever. That particular host cell no longer can be affected by the parasites and neither can its descendants. So the mutant hosts can breed happily while the other hosts, not immune to the parasites, get killed off.
Victory? Well, if the parasites were to all stay as they are, then they'd eventually die out or go infect something else when the infectable hosts are all killed off. But what's more likely to happen is that one of the parasites suffers a mutation that allows it to infect 'immune' hosts too. So while the non-mutated parasites die off from starvation, mutant parasites proliferate on mutant hosts and we're back at square one, in a typical evolutionary arms-race in which the stakes are life and death.
Which is indeed the typical situation. However, now imagine this. One of the hosts suffers a mutation which, while it doesn't make it immune to the parasite infestation, it prevents the parasites to multiply once inside that host. Or rather, it prevents them from multiplying apart from the moment that the host divides itself into two descendants. If you like, imagine that the host can keep a 'cap' on how many parasites live in it. If it splits into two, the two new hosts can each support a few more parasites, allowing those inside to breed as well.
The stakes of the arms-race have changed. They are still life and death for the host, but for the parasites the stake is now reproductive freedom. It's a subtle change, but one that makes all the difference.
For, if the parasites find no mutation to counteract the control imposed by the host, they nonetheless do not die out. So descendants of parasites that don't fight that control will be around.
Of course, if a parasite stumbles into a mutation that frees it, we're back in the typical arms race. Which, for the parasite, is a very good strategy to get more descendants.
But there's another way for a parasite under the reproductive control of the host to get more descendants. It is to encourage and enable the host to reproduce more often. If one of the parasites stumbles into a mutation that does that, it now actually improves the fitness of the host instead of diminishing it.
That is the moment when parasite becomes symbiont.
My guess is that remaining parasitic is the best way to get the most descendants, so most of the descendants of the original parasite will still be parasites in to the death arms races with the hosts. However, becoming a symbiont is a decent enough strategy, so there'll be a good number of them too. Finally, a dwindling part of the population of parasites will be the 'lazy' ones, that don't either fight the control nor improve the host.
Let's now focus on the host and symbiont population, because something interesting is happening here. The context for the two organisms has changed, meaning the deciding factors of what makes a mutation deleterious or beneficial have also changed.
If a mutation made a parasite's membrane leaky it would result in it spilling its 'guts' and dying, if it were to also live outside a host. Such a mutation would be deadly and deleterious for a parasite. But a symbiont living inside the cell might not suffer because of a leaky membrane. In fact, it may be improved by it. So while in parasites a leaky membrane mutation would disappear, this would not be the case for symbionts.
Likewise, a mutation which made the host unable to process an important nutrient would starve and kill it. Deleterious, unless the host already has some symbionts capable to process that nutrient for it, perhaps more even more efficiently now that the host has given up that task to the symbiont. In that case, the host can live and get descendants who also rely on symbionts.
These co-dependency mutations are not just side effects, they serve the important purpose of 'sealing the deal'. For, if a leaky symbiont suddenly mutates to break the host's control, it will kill it, but then it would itself die and its mutation doesn't get a future generation. Similarly, a host that depends on its symbionts and who mutates to get an immune response that kills them will also die. That mutation won't see a future either. Therefore, mutations that would have been beneficial, if the host and symbiont were locked in a to the death arms race, are now deleterious. It is unlikely that either side will break the partnership and live to spread its cheating genes.
The above is a "just so story" because it relies on mutations to do some fairly specific things, in particular reproductive control over the parasites. Are such mutations even possible? The answer appears to be yes, but this was not found by theorizing in just so stories. It was found by looking at what goes on inside a cell. Mitochondria, chloroplasts and other structures suspected to have started as possibly parasitic bacteria reproduce in synchrony with the larger cell. They cannot live outside that cell, neither can that cell live without them. And by the nature of how the most basic of organisms, the common ancestors of plants and animals today, reproduced, that whole story of mutations that I've imagined above only needed to happen once. Well, once per co-opted bacteria, but still. One host and one symbiont are enough to be the common ancestor of all plants that use chloroplasts. One host and one symbiont are enough to be the common ancestor of all animals. So that story I told is quite plausible- and I'd settle for nothing less of course!
Whatever the exact mechanism of its construction- the synchronization of reproduction events between host and symbionts appears to be a key element-, an endosymbiotic partnership may be the oldest alliance that I know of in the universe. We and our distant ancestors have relied on our mitochondria to power our cells for hundreds of millions of years. That's an unimaginably long span of time. Mitochondria are so deeply integrated that they are an unquestioned and unquestioningly loyal part of us, even as their (and our) distant cousins, the bacteria that stayed in the arms race, are plaguing us with disease.
I'll end this post on another note of weirdness. Caution, the following isn't yet proven in humans, but it appears to be established for mice, is plausible for all mammals including us, and has a certain poetic resonance. The placenta, the organ that surrounds the growing foetus inside the mother, contains some strange, three-nuclei cells, their purpose being to make sure that the growing baby is not attacked and destroyed by the immune system of the mother. It appears that the proteins that these mediator cells use, syncytins, are produced by genes that the ancestor of mammals had absorbed from something like an immunodeficiency virus.
So yeah. The fact that we can be carried in a womb and then be born alive is the result of coopting a distant cousin of HIV. Put that in your pipe and smoke it.
Since Prometheus the film deals with lofty things as the origin of life and whatnot, I thought endosymbiosis is a worthy topic to try and reference, however oblique and distant the reference ends up in the text. For one, it neatly contrasts with the view of life as eternal, deadly competition. That's a true view, by the way; the circle of life isn't nearly as nice as new age sentiment makes it to be. Life is vicious, cruel, short-sighted and selfish. Paradoxically, this blind idiot monster sometimes produces incredibly long-lasting partnerships, and trying to explain how that may happen is the point of this post. (In the fic, the endosymbiosis is engineered by intelligent agents, the main characters. For the real life endosymbiotic partnerships, to the best available evidence, there was no such thing.)
Another reason why endosymbiosis is interesting, to me, is that it's a genuinely cool, genuinely mad idea that, unlike most mad ideas in science, withstood the test of evidence and came out the stronger for it. You might think, if you went by the crap that passes for pop-sci these days, that crazy and worthy ideas happen all the time: relativity, quantum mechanics, quantum electrodynamics, chaos theory and so on. False. Those first three were built on a solid foundation of experiments that older theories couldn't explain. The last (chaos theory) was simply the natural consequence of classical mechanics as revealed by the high powered mathematics emerging at the end of the nineteenth century. All of that may be fodder for other posts though.
No, a better example of a similarly crazy and worthy idea is continental drift. When Alfred Wegener proposed that continents slowly move across the surface of the Earth, he really had nothing but a hunch based on the similarity of the coasts of South America and Africa. When Konstantin Mereschkowski and Andreas Schimper proposed that chloroplasts, tiny structures in plant cells that are responsible for photosynthesis, were originally independent bacteria, they really only had a hunch based on the fact that chloroplasts kinda looked like bacteria.
 |
| Chloroplasts in leaf cells of Plagiomnium Affine. Image taken from wikipedia where it is posted under the GNU Free Documentation licence. |
That was scant evidence, at best. It so happens however that in these two cases the hunches turned out to be true. To the best of our knowledge today, chloroplasts indeed descend from cyanobacteria that were, in some distant past, independent of the plant cell. One aspect of that proof is that they have their own DNA with great similarities to that of cyanobacteria. Mitochondria, structures in both plant and animal cells that produce ATP to fuel the cells' other activities, also used to be independent bacteria eons ago. They carry their own DNA too, and since only the egg cell transports mitochondria into a foetus, using mitochondrial DNA one can trace the maternal lineage of somebody.
 |
| Mitochondria in mammalian lung cell as seen under an electron microscope. Image taken from wikipedia where it is posted as public domain by the image's author, Louisa Howard. |
Now, a disclaimer. I'm not a biologist. I will try to give an honest, clear and concise summary of what I understand about evolution and endosymbiosis though, even if all I can produce is, in fairness, a just-so story. Still, I think I'm not too far off in how I imagine the thing to work. So onwards, beginning with a brief summary on evolution.

First, mutations. In the figure I tried to represent how mutations spread in a population of bacteria that reproduce asexually. Each bacteria will, if it lives long enough, split itself into two smaller bacteria at some point, which will grow to normal size using nutrients from their surroundings. Initially there's one 'blue' bacteria, and I'll often refer to all of its descendents together as a 'population'.
Even if at some point not all descendents are alike. In the figure, the original bacteria splits into two, and these two split again to make four, but a mutation occurs and now one of the bacteria is 'red'. Next generation, the blue bacteria split into more blue bacteria, while the red is now the ancestor of a small population of red bacteria. Both versions, 'blue' and 'red' may coexist in the population of descendants of the original blue.
Sometimes a mutation is so harmful that it prevents the organism it appears in from reproducing. Such a mutation is deleterious, like the 'green' in the figure. Since green doesn't live long enough to get descendants, that mutation disappears.
Most mutations are neutral. In the figure, 'red' and 'blue' are more or less doing equally well. Context, that is, the environment that organisms live in, is what will select among mutations. Some mutations will make the organism that carries them better able to get descendants in the environment it lives in. In the figure, 'purple' is especially prolific, and will tend to dominate the population of descendants of the original blue.
Note, this doesn't necessarily mean that blue and red disappear. Unless there's something in the environment that actively kills them off, they'll stick around. In lesser numbers than purple, true. But evolution is NOT about being the most efficient, nor the most prolific. As long as you can get a future generation, you're doing well enough (of course, being prolific and efficient helps).
With that basis set up, let's tackle endosymbiosis. For this, imagine the following scenario. Suppose there's a population of single celled animals like amoebas or something similar, merrily going about their single celled lives. These will be the 'hosts'. Now, there are also, floating around, some parasitic bacteria that have the nasty habit of infiltrating inside a host cell, feasting on its contents and multiplying prodigiously, killing it in the process. These will be the 'parasites'.

So the parasites cause a deadly disease in a host, if they infect it. So far, so fine. Evolution in a typical host-parasite competition. Because, suppose one of the host cells suffers a mutation that makes it immune to the parasite infection. They can't get in or are killed off if they do, whatever. That particular host cell no longer can be affected by the parasites and neither can its descendants. So the mutant hosts can breed happily while the other hosts, not immune to the parasites, get killed off.
Victory? Well, if the parasites were to all stay as they are, then they'd eventually die out or go infect something else when the infectable hosts are all killed off. But what's more likely to happen is that one of the parasites suffers a mutation that allows it to infect 'immune' hosts too. So while the non-mutated parasites die off from starvation, mutant parasites proliferate on mutant hosts and we're back at square one, in a typical evolutionary arms-race in which the stakes are life and death.
Which is indeed the typical situation. However, now imagine this. One of the hosts suffers a mutation which, while it doesn't make it immune to the parasite infestation, it prevents the parasites to multiply once inside that host. Or rather, it prevents them from multiplying apart from the moment that the host divides itself into two descendants. If you like, imagine that the host can keep a 'cap' on how many parasites live in it. If it splits into two, the two new hosts can each support a few more parasites, allowing those inside to breed as well.
The stakes of the arms-race have changed. They are still life and death for the host, but for the parasites the stake is now reproductive freedom. It's a subtle change, but one that makes all the difference.
For, if the parasites find no mutation to counteract the control imposed by the host, they nonetheless do not die out. So descendants of parasites that don't fight that control will be around.
Of course, if a parasite stumbles into a mutation that frees it, we're back in the typical arms race. Which, for the parasite, is a very good strategy to get more descendants.
But there's another way for a parasite under the reproductive control of the host to get more descendants. It is to encourage and enable the host to reproduce more often. If one of the parasites stumbles into a mutation that does that, it now actually improves the fitness of the host instead of diminishing it.
That is the moment when parasite becomes symbiont.
My guess is that remaining parasitic is the best way to get the most descendants, so most of the descendants of the original parasite will still be parasites in to the death arms races with the hosts. However, becoming a symbiont is a decent enough strategy, so there'll be a good number of them too. Finally, a dwindling part of the population of parasites will be the 'lazy' ones, that don't either fight the control nor improve the host.
Let's now focus on the host and symbiont population, because something interesting is happening here. The context for the two organisms has changed, meaning the deciding factors of what makes a mutation deleterious or beneficial have also changed.
If a mutation made a parasite's membrane leaky it would result in it spilling its 'guts' and dying, if it were to also live outside a host. Such a mutation would be deadly and deleterious for a parasite. But a symbiont living inside the cell might not suffer because of a leaky membrane. In fact, it may be improved by it. So while in parasites a leaky membrane mutation would disappear, this would not be the case for symbionts.
Likewise, a mutation which made the host unable to process an important nutrient would starve and kill it. Deleterious, unless the host already has some symbionts capable to process that nutrient for it, perhaps more even more efficiently now that the host has given up that task to the symbiont. In that case, the host can live and get descendants who also rely on symbionts.
These co-dependency mutations are not just side effects, they serve the important purpose of 'sealing the deal'. For, if a leaky symbiont suddenly mutates to break the host's control, it will kill it, but then it would itself die and its mutation doesn't get a future generation. Similarly, a host that depends on its symbionts and who mutates to get an immune response that kills them will also die. That mutation won't see a future either. Therefore, mutations that would have been beneficial, if the host and symbiont were locked in a to the death arms race, are now deleterious. It is unlikely that either side will break the partnership and live to spread its cheating genes.
The above is a "just so story" because it relies on mutations to do some fairly specific things, in particular reproductive control over the parasites. Are such mutations even possible? The answer appears to be yes, but this was not found by theorizing in just so stories. It was found by looking at what goes on inside a cell. Mitochondria, chloroplasts and other structures suspected to have started as possibly parasitic bacteria reproduce in synchrony with the larger cell. They cannot live outside that cell, neither can that cell live without them. And by the nature of how the most basic of organisms, the common ancestors of plants and animals today, reproduced, that whole story of mutations that I've imagined above only needed to happen once. Well, once per co-opted bacteria, but still. One host and one symbiont are enough to be the common ancestor of all plants that use chloroplasts. One host and one symbiont are enough to be the common ancestor of all animals. So that story I told is quite plausible- and I'd settle for nothing less of course!
Whatever the exact mechanism of its construction- the synchronization of reproduction events between host and symbionts appears to be a key element-, an endosymbiotic partnership may be the oldest alliance that I know of in the universe. We and our distant ancestors have relied on our mitochondria to power our cells for hundreds of millions of years. That's an unimaginably long span of time. Mitochondria are so deeply integrated that they are an unquestioned and unquestioningly loyal part of us, even as their (and our) distant cousins, the bacteria that stayed in the arms race, are plaguing us with disease.
I'll end this post on another note of weirdness. Caution, the following isn't yet proven in humans, but it appears to be established for mice, is plausible for all mammals including us, and has a certain poetic resonance. The placenta, the organ that surrounds the growing foetus inside the mother, contains some strange, three-nuclei cells, their purpose being to make sure that the growing baby is not attacked and destroyed by the immune system of the mother. It appears that the proteins that these mediator cells use, syncytins, are produced by genes that the ancestor of mammals had absorbed from something like an immunodeficiency virus.
So yeah. The fact that we can be carried in a womb and then be born alive is the result of coopting a distant cousin of HIV. Put that in your pipe and smoke it.
Comments
Post a Comment